
Recently, I showed that a simple genetic algorithm with uniform crossover can solve a noisy learning 7-parity problem efficiently—in 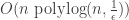

The implicit concurrency hypothesis posits that the efficient learning of relevant variables in the presence of large numbers of irrelevant variables is precisely what gives evolution its power; so, understandably, when I read about the learning juntas problem a few weeks ago (on this blog), I sensed the possibility of a connection. Formally, a 




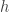
Observe how this problem abstracts the genetic epidemiology problem mentioned above. The junta are the genetic loci participating in the presence/absence of the disease. The hidden function specifies which configurations of alleles at these loci cause the disease, and which ones do not. The problem of learning the relevant loci is the problem of learning juntas. Also note that the learning juntas problem generalizes the learning parities problem straightforwardly. Whereas the hidden function in the case of learning parities must be the parity function, the hidden function in the case of learning juntas can be any boolean function over the juntas.
Here is an evolution based algorithm, written in Python, that learns juntas given a membership query oracle. And here is a tutorial on its usage. For any value of 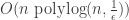
Implications for the Neo-Darwinian Synthesis
It seems like more than a coincidence that
- The Learning Juntas problem abstracts the genetic epidemiology problem mentioned earlier
- Evolutionary computation can efficiently solve the Learning Juntas problem for small numbers of juntas.
One is hard pressed not to hypothesize that evolution takes the form it takes precisely because this form allows it to solve a problem that is similar to the genetic epidemiology problem in all significant ways, save two :
- The problem is to learn the genetic loci participating in fitness/hotness not disease penetrance (often the same loci, but by no means always)
- Unlike computational geneticists, evolution has access to an active oracle.
Given the straightforwardness of this idea it may seem odd that it has not been forthcoming from Evolutionary Biology itself; that is until one realizes that a deep-rooted practice in Population Genetics actively steers theorists away. A vestige from the days before computers, when pen, paper, and mathematics was all one had, the conflation of the unit of inheritance with the unit of selection casts a long shadow. More about this in Sections 2.4.1 and 5.1 of my dissertation.
What now?
Despite the routine use of evolutionary algorithms in science and engineering, computer scientists have been unable to give a straight answer to a simple question: “Is there anything computationally efficient about evolution?” This question can now be answered in the affirmative. Recombinative evolutionary algorithms are capable of efficiently solving a broad, non-trivial problem: Learning
While fascinating from a biological perspective, this answer is not entirely satisfying to the engineer in me because there are conventional algorithms that solve the problem under consideration as efficiently; for example, the binary search algorithm due to Blum and Langley described in Section 5 of the paper by Mossel et al. A more involved algorithm due to Feldman solves the problem almost as efficiently, and does so non-adaptively and in the presence of random persistent classification error.
Have modern ML techniques eclipsed evolution at the very thing it is good at?
I have a hunch that the answer is “no”. It would be fantastic to find generalizations of the learning juntas with membership queries problem that are more difficult for conventional algorithms, but remain efficiently solvable by evolution.
Relevant Computational Learning Theory papers
R.J. Lipton
[web] The Learning Juntas Problem
(An excellent introduction to the Learning Juntas problem)
E. Mossel, R. O’donnell, and R. Servedio
[pdf] Learning Juntas,
(Section 3.1 is especially relevant if you want to reverse engineering the posted Python algorithm)
A. Blum, L. Hellerstein, and N. Littlestone
[pdf] Learning in the Presence of Finitely or Infitely Many Irrelevant Attributes
V. Feldman
[pdf] Attribute-Efficient and Non-adaptive Learning of Parities and DNF Expressions
Relevant papers on Implicit Concurrency
K.M. Burjorjee
[pdf] The Fundamental Learning Problem that Genetic Algorithms with Uniform Crossover Solve Efficiently and Repeatedly as Evolution Proceeds
K.M. Burjorjee
[pdf] Explaining Optimization in Genetic Algorithms with Uniform Crossover
Relevant papers on genetic epidemiology
J.H. Moore, L.W. Hahn, M.D. Ritchie, T.A. Thornton, and B.C. White
[web] Routine Discovery of Complex Genetic Models using Genetic Algorithms
J.H. Moore
[web] The ubiquitous nature of epistasis in determining susceptibility to common human diseases
Last Updated: September 18, 2013

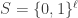 be a search space consisting of binary strings of length
be a search space consisting of binary strings of length  . Let
. Let 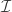 be some set of indices between
be some set of indices between 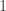 and
and 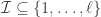 . Then
. Then  into
into 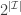 subsets called schemata (singular schema) as in the following example: suppose
subsets called schemata (singular schema) as in the following example: suppose 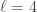 , and
, and  , then
, then 


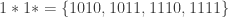
 stands for ‘wildcard’. Partitions of this type are called schema partitions. As we’ve already seen, schemata can be expressed using templates, for example,
stands for ‘wildcard’. Partitions of this type are called schema partitions. As we’ve already seen, schemata can be expressed using templates, for example,  . The same goes for schema partitions. For example
. The same goes for schema partitions. For example  denotes the schema partition represented by the index set
denotes the schema partition represented by the index set 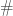 stands for ‘defined bit’. The order of a schema partition is simply the cardinality of the index set that defines the partition (in our running example, it is
stands for ‘defined bit’. The order of a schema partition is simply the cardinality of the index set that defines the partition (in our running example, it is  ). Clearly, schema partitions of lower order are coarser than schema partitions of higher order.
). Clearly, schema partitions of lower order are coarser than schema partitions of higher order. is
is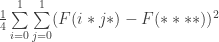
 gives the average fitness of a schema under sampling from the uniform distribution.
gives the average fitness of a schema under sampling from the uniform distribution.![[\mathcal I]](https://s0.wp.com/latex.php?latex=%5B%5Cmathcal+I%5D&bg=ffffff&fg=333333&s=0&c=20201002) denote the schema partition represented by some index set
denote the schema partition represented by some index set  , and let
, and let  . Then
. Then  is
is  . So for
. So for  , the number of schema partitions of order 2,3,4 and 5 are on the order of
, the number of schema partitions of order 2,3,4 and 5 are on the order of  , and
, and  respectively. The take away from our thought experiment is this: while a search space may have vast numbers of coarse schema partitions, most of them will have negligible effects (due to averaging). In other words, while coarse schema partitions are numerous, ones with non-negligible effects are rare.
respectively. The take away from our thought experiment is this: while a search space may have vast numbers of coarse schema partitions, most of them will have negligible effects (due to averaging). In other words, while coarse schema partitions are numerous, ones with non-negligible effects are rare. combinations of four indices between between 1 and 200. Given some bitstring, if the parity of the bits at these indices is 1 (i.e. if the sum of the four bits is odd) then the stochastic function returns a value drawn from the magenta distribution (see below). Otherwise, it returns a value drawn from the black distribution. The four indices are said to be pivotal. All other indices are said to be non-pivotal.
combinations of four indices between between 1 and 200. Given some bitstring, if the parity of the bits at these indices is 1 (i.e. if the sum of the four bits is odd) then the stochastic function returns a value drawn from the magenta distribution (see below). Otherwise, it returns a value drawn from the black distribution. The four indices are said to be pivotal. All other indices are said to be non-pivotal.
 possible combinations of four loci. From all of these possibilities, the genetic algorithm managed to identify the correct one within five hundred generations.
possible combinations of four loci. From all of these possibilities, the genetic algorithm managed to identify the correct one within five hundred generations. , the genetic algorithm solves the problem with no increase in the number of queries. Not bad. (Of course we’re talking about a single run of a stochastic process. And yes, it’s a representative run. See chapter 3 of my
, the genetic algorithm solves the problem with no increase in the number of queries. Not bad. (Of course we’re talking about a single run of a stochastic process. And yes, it’s a representative run. See chapter 3 of my  combinations of four loci. That’s on the order of a million trillion combinations. This time round, let’s also change the locations of the 4 pivotal loci. Will the genetic algorithm find them in 500 generations or less?
combinations of four loci. That’s on the order of a million trillion combinations. This time round, let’s also change the locations of the 4 pivotal loci. Will the genetic algorithm find them in 500 generations or less?